
0371-60972711
來源: 點擊:1800 喜歡:0
2016-11-21 10:10:26 來源: 點擊:
污水活性污泥處理過程中會產生大量的剩余污泥,數量可達到污水處理量的0.3%~0.5%(以含水率97%計)[1].剩余污泥除了具有含水率高、 易腐爛、 惡臭等特征外,還含有大量的病原菌、 寄生蟲、 重金屬和二 英、 苯并芘等難以降解的有毒、 有害、 致癌物質,極易對土壤、 地下水等造成二次污染[2].厭氧消化處理是對剩余污泥進行穩定化、 減量化和資源化過程中被廣泛采用的處理手段,具有能耗低、 污泥穩定性好、 產生生物能源沼氣等優點[3].影響剩余污泥厭氧消化過程的因子包括基礎因素(厭氧污泥組成、 濃度、 污泥負荷等)和環境因素(pH、 ORP、 抑制性物質等)兩大類,其中厭氧污泥的生物相組成和代謝活性對厭氧消化處理的過程進展發揮著重要的作用[4].在剩余污泥厭氧消化過程中,由于微生物構成、 對基質的適應性和接種量的不同,采用不同的接種厭氧污泥會對剩余污泥產CH4生成勢形成不同程度的影響[5,6].深入探究剩余污泥厭氧消化過程中產CH4生成勢與菌群動態變化的關系,一方面可對厭氧消化過程中剩余污泥的生化降解過程和產CH4潛能進行評價[7],另一方面也能為剩余污泥厭氧消化工藝的關鍵操作參數優化提供依據[8,9]剩余污泥厭氧消化的效率在很大程度上取決于厭氧微生物種群多樣性及優勢種群的活性[10,11].不同條件下厭氧消化運行的穩定性及效率與系統群落結構的變遷會存在一定的關聯.厭氧污泥中主要存在水解發酵菌、 產氫產乙酸菌、 產甲烷菌及硫酸鹽還原菌[12].其中產甲烷菌屬于典型的古細菌,大致可以分為兩類:一類主要利用乙酸產生甲烷,主要有產甲烷八疊球菌(Methanosarcina)和產甲烷髦毛菌(Methanothrix); 另一類利用氫和二氧化碳合成甲烷.由于傳統微生物培養、 鑒定的局限性,近年來研究人員嘗試應用基于16S rRNA的分子生物學技術(變性梯度凝膠電泳、 克隆文庫技術、 熒光原位雜交)對厭氧污泥系統群落結構的變化進行分析.其中末端限制性片段多態性(terminal-restriction fragment length polymorphism, T-RFLP)根據PCR擴增產物片斷的大小不同以及標記片斷種類和數量的不同來分析群落的結構及組成. Collins等[13]利用T-RFLP技術對接種污泥和接種后的污泥中微生物菌群變化進行研究后發現接種污泥中占優勢的產甲烷菌群是Methanosarcinales、 Methanobacteria和Proteobacteria,而反應器運行穩定后占優勢的菌群為Methanosarcina vacuolata和Methanobacterium palustre.T-RFLP技術可以很靈敏地檢測微生物種類的微小變化,能夠提供微生物種群結構和數量動態變化的信息,已成功應用于厭氧污泥產CH4菌的群落結構、 動態變化的檢測等方面[14].
本研究采用剩余污泥厭氧消化產CH4生成勢(biological methane potential,BMP)的測試方法[15, 16],對兩廠的剩余污泥厭氧消化進行了批次實驗,在兩廠剩余污泥的產的產CH4速率、 基質濃度回歸的基礎上得出產CH4的關鍵參數,評價不同剩余污泥在厭氧消化過程中的產CH4生成勢; 同時對BMP實驗前后的水質變化進行分析,結合厭氧消化前后T-RFLP的變化,對兩種剩余污泥在厭氧消化過程中CH4生成勢的差異進行解析[17].一方面可對不同剩余污泥厭氧消化過程中的產CH4潛能進行評價,同時也能為厭氧消化工藝中微生物菌群動態變化跟蹤及關鍵參數優化提供依據.
1 材料與方法
1.1 厭氧污泥來源與特征
本研究所用的剩余污泥、 厭氧污泥分別來自于AP和DH污水處理廠,AP污水處理廠采用合流制明渠排水溝進水,進水水質易受降雨影響,且泥沙等無機顆粒含量較高; DH污水處理廠配套管網設施完善,進水為完整的下水管網收集的城市生活污水.污水廠長期的監測數據表明,AP厭氧污泥的VSS/TSS值多在0.5以下,而DH厭氧污泥的VSS/TSS值維持在0.6以上,厭氧消化池代謝活性較好.AP剩余污泥的VSS為8 050 mg ·L-1,TSS為14 350 mg ·L-1.DH剩余污泥的VSS為9 250 mg ·L-1,TSS為13 230 mg ·L-1. 1.2 BMP實驗設計
AP-BMP、 DH-BMP共設置6組不同的污泥負荷F/M(0、 0.1、 0.25、 0.4、 0.6、 1)進行實驗,由于污泥濃縮的不均衡性,實測F/M如表1所示.BMP實驗在120 mL的血清瓶中進行,依據測試基質濃度加入定量剩余污泥.采用Owen等[15]提出的厭氧微量元素溶液配方,其中CaCl2 ·2H2 O、 NH4Cl、 MgCl2 ·6H2 O、 KCl、 MnCl2 ·4H2 O、 CoCl2 ·6H2 O、 H3BO3、 CuCl2 ·2H2 O、 Na2MoO4 ·2H2 O、 ZnCl2的濃度分別為16.7、 26.6、 120、 86.7、 1.33、 2、 0.38、 0.18、 0.17、 0.14 g ·L-1.每個血清瓶加入27 mL的微量元素液體和5.4 mL的(NH4)2HPO4(26.7 g ·L-1),之后依據不同的F/M比值接種厭氧污泥、 剩余污泥后,蓋上膠蓋并用鋁箔封口.在35℃下旋轉培養箱內進行實驗,實驗過程中以玻璃注射筒(50 mL)測量總產氣量,并利用GC-ECD測定CH4、 CO2和H2的比例.共設置兩組平行實驗,取均值進行產氣量、 水質分析. AP-BMP、 DH-BMP分別設置兩組平行實驗,產氣量測量可精確到0.1 mL.
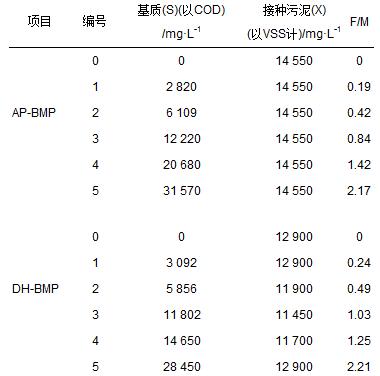
表 1 BMP實驗實測F/M比
1.3 分析方法
1.3.1 常規指標
常規水質指標,包括pH(Suntex Ion Analyzer 3000A)、 TSS(Sartorius Analytic oven)、 NH+4-N(Autotitrator AT-400KYOTO)、 TKN(Autotitrator AT-400KYOTO)、 COD(回流加熱滴定)等,均依照美國EPA規定的Standard Methods[18]規定進行.實驗過程中氣體成分(CH4、 CO2和H2)的測量采用氣相色譜(China Chromatography GC8900T)進行.
1.3.2 T-RFLP分析
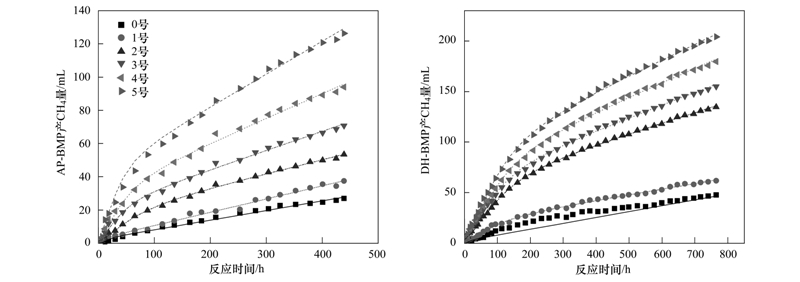
圖1 實測CH4累積產氣量與回歸曲線
本研究參照了Lueders等[19]建立的T-RFLP方法對實驗前后厭氧污泥的生物多樣性進行分析,實驗步驟如下:①通過PCR來復制DNA樣品中的目標基因,引物為在5′的尾端上帶有熒光的Ar 109f和Ar 912r*,首先在94℃預變性5 min,擴增循環階段包括94℃變性1 min,52℃退火1 min,72℃延伸1.5 min,共循環28 cycles,最后在72℃條件下延伸6 min.②在8.5 μL的PCR產物中加入0.5 μL TaqⅠ和1.5 μL的緩沖溶液,在65℃條件下消化切割2 h; ③將切割后的片段利用電泳分離并以熒光偵測器(全自動遺傳分析儀,ABI PRISM 3100 Genetic Analyzer)檢測片段上所帶的熒光強度. Lueders等[19]成功分離、 克隆出產CH4髦毛菌Methanosaeta spp.、 產CH4微菌Methanomicrobiaceae、 RC-I和產CH4桿菌Methanobacteriaceae,未能定性的菌種歸入Diverse類.
1.3.3 數據分析
剩余污泥厭氧消化過過程中累積CH4產氣量采用改進的Gompertz模型回歸分析[20]

式中,y為累積產氣量(mL); δ為產氣末期校正斜率(mL·h-1); t為反應時間(h); A為平衡產氣量(mL); Rmax:最大產氣速率(mL ·h-1); λ:遲滯期(h). 底物的厭氧代謝過程實質上是一系列的酶促反應,因此采用Michaelis-Menten模型描述剩余污泥濃度與比產氣速率的關系: V=Vmax · S Km+S (2) 式中,Vmax為最大比產氣速率[mL ·(g ·d)-1]; V為比產氣速率[mL ·(g ·d)-1]; Km為半飽和常數(mg ·L-1); S為基質濃度(mg ·L-1).
2 結果與討論
2.1 BMP產氣結果分析
AP-BMP、 DH-BMP批次實驗持續時間分別為439 h、 765 h,6組不同的投配比設計累積產氣量存在著明顯的差異.實驗初期累積產氣量差異不明顯(圖1),后期逐步增大.兩組實驗氣體成分均以CH4(80%)、 CO2(20%)為主,同時存在少量的H2(不足1%).實驗產氣均勻,可分為兩個階段.第一階段產氣速率大,產甲烷菌利用溶解性COD或易水解性物質進行厭氧發酵; 之后產氣速率逐漸降低至穩定水平,該階段的限速步驟為剩余污泥的水解過程,產氣隨著水解的進程而穩定增加.至該實驗結束時,每天仍有穩定體積的氣體產生,表明微生物的水解、 厭氧產CH4仍在進行中.應用改進的Gompertz模型對AP-BMP、 DH-BMP產氣進行回歸后的參數如表2所示.兩廠的厭氧污泥產氣數據與改進的Gompertz模型擬合較好(R2>0.99).通常厭氧消化過程中的限速步驟為水解破壁過程[21, 22],在無添加外來基質的0號樣品中,AP、 DH厭氧污泥的遲滯期分別為20.00 h、 12.93 h,表明DH厭氧污泥的活性較高.

表2 改進Gompertz模型回歸BMP實驗參數

圖2 Michaelis-Menten模型回歸BMP產CH4速率參數
采樣Michaelis-Menten模型對兩個批次實驗的基質濃度、 比產氣速率回歸后的結果如圖2所示(R2>0.99).AP、 DH厭氧污泥的最大比產氣速率差距不大,分別為74.21、 51.99 mL ·(g ·d)-1,但兩廠厭氧污泥的Km存在著顯著的差異,DH厭氧污泥的Km為19 005 mg ·L-1而AP厭氧污泥的Km高達54 098 mg ·L-1.Km是表征底物親和力的常數,表明AP厭氧污泥對該廠剩余污泥的適應性差,需要在較高的基質濃度下才能表現出較好的產CH4性能.
2.2 水質變化分析
AP-BMP、 DH-BMP兩個批次實驗結束時不同F/M條件下的水質狀況如下表3所示.兩個批次實驗中隨著F/M增大,pH由弱酸性漸變為弱堿性,分別位于在6.8~7.2、 6.7~7.1范圍內.0號呈弱酸性,可能由未添加基質,厭氧污泥自身的水解造成[23].DH-BMP與AP-BMP實驗結束時ORP值分別位于-235~-280 mV與-235~-282 mV范圍內.隨著F/M比的增大,兩個批次實驗的ORP值都呈下降的趨勢.厭氧環境的主要標志是發酵液具有低的ORP,不同的厭氧消化體系和不同的厭氧微生物對ORP的要求不同[24].
兩批次實驗中在高F/M條件下下CODT下降明顯.與AP-BMP實驗相比,DH-BMP實驗結束后CODT下降更為顯著,5號樣品中由之前的47 000 mg ·L-1下降至32 480 mg ·L-1,在高F/M條件下厭氧消化進行得更為完全.批次實驗前后,NH+4-N濃度均有提高,不同F/M條件下NH+4-N變化差異較大,DH-BMP實驗中5號樣品實驗結束后NH+4-N濃度達到了755.5 mg ·L-1.污泥厭氧消化過程中,污泥的水解是限速步驟.在高F/M條件下,厭氧污泥代謝旺盛,使得剩余污泥的破壁、 氨化過程進行得較為完全,從而使得NH+4-N顯著提升[4].兩個批次實驗前后TSS變化呈現出與CODT類似的趨勢,高F/M條件下下降趨勢明顯,DH-BMP實驗中5號樣品的TSS由實驗前的45 100 mg ·L-1下降至38 050 mg ·L-1.綜合比較AP-BMP與DH-BMP實驗前后水質變化,DH-BMP實驗前后CODT、 TSS下降趨勢更加明顯,表明DH-BMP過程中代謝活躍,厭氧消化過程進行得較為完全.


表3 BMP實驗末期不同F/M條件下主要水質
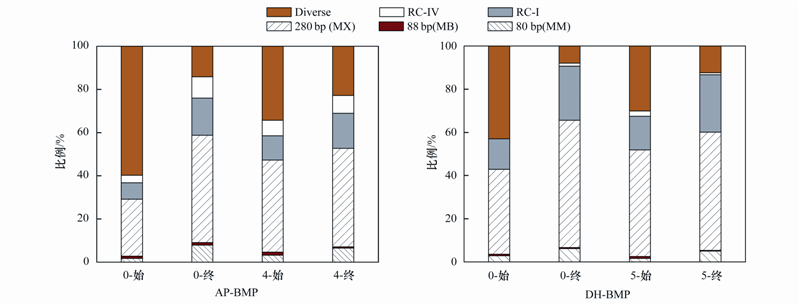
圖3 AP、 DH剩余污泥厭氧消化前后主要T-RFs豐度變化
2.3 厭氧污泥生物多樣性分析
圖3表示了以AP-BMP、 DH-BMP實驗前后產甲烷功能菌組成的變化情況.與AP-BMP、 DH-BMP兩個批次實驗中產CH4、 水質變化情況相一致,DH接種厭氧污泥中產甲烷菌群的相對含量高于AP接種厭氧污泥,AP-BMP中0號樣品的雜菌(Diverse)相對含量超過60%.DH厭氧污泥中產甲烷功能菌群豐富且相對含量較高,可能是DH-BMP實驗中半飽和常數Km顯著低于AP-BMP中回歸數值的重要原因.
AP-BMP實驗后,未添加基質的0號實驗結束后雜菌(Diverse)相對含量降至14%,這可能是由于基質的缺乏,產甲烷菌將雜菌(Diverse)分解代謝造成.4號樣品中的雜菌(Diverse)含量也有所降低但幅度不大.同時實驗結束后,兩組的產甲烷髦毛菌Methanosaeta spp.(280 bps)、 產甲烷微菌Methanomicrobiaceae(80 bps)和RC-I(389 bps)的相對含量皆有提高.DH-BMP實驗前后產甲烷功能菌的組成變化情況與AP-BMP變化類似.DH-BMP實驗結束時雜菌(Diverse)含量降低,0號、 5號樣品中雜菌(Diverse)相對含量不足17%.產甲烷髦毛菌Methanosaeta spp.(280 bps)、 產甲烷微菌Methanomicrobiaceae (80 bps)和RC-I(389 bps)的相對含量皆有所提高,表明產甲烷菌的活性和相對豐度在實驗過程中得以提高.已有研究發現產甲烷髦毛菌Methanosaeta spp.主要以乙酸為基質生成甲烷[25],實驗結束后產甲烷髦毛菌Methanosaeta spp.相對含量提高,而其它兩種產甲烷菌以H2和CO2為基質生成甲烷,由于兩組BMP實驗中H2的生成量較低(不足1%),因此產甲烷桿菌Methanobacteriacea (88 bps)、 產甲烷微菌Methanomicrobiaceae(80 bps)相對含量變化不如產甲烷髦毛菌Methanosaeta spp.(280 bps)增加顯著。
3 結論
(1)AP、 DH剩余污泥厭氧消化產CH4采用改進Gompertz模型回歸的最大比產氣速率數值接近,分別達到了74.21 mL ·(g ·d)-1、 51.99 mL ·(g ·d)-1,但對基質剩余污泥的半飽和常數Km差異較大,分別為54 098 mg ·L-1和19 005 mg ·L-1,表明AP剩余污泥親和性差、 難于厭氧消化.
(2)BMP實驗結束后,TSS、 CODT有所下降,NH+4-N顯著提高,且高污泥負荷F/M條件下水質變化趨勢更為顯著.與AP-BMP實驗相比,DH-BMP水質變化顯著.
(3)兩批次實驗前后T-RFLP分析結果與其生化產CH4勢相一致,實驗結束后雜菌(Diverse)相對含量顯著下降,產甲烷髦毛菌Methanosaeta spp.(280 bps)、 產甲烷微菌 Methanomicrobiaceae(80 bps)和RC-I(389 bps)的相對含量皆有所提高,且DH-BMP樣品中產甲烷功能菌群相對含量提升程度高于AP-BMP.

